

兩棲綱及爬行綱 (AMPHIBIA et REPTILIA)
兩棲綱 Class: AMPHIBIA
殼椎亞綱 Subclass: Lepospondyli
(1) 游螈目 Nectridea

盜首螈 Diplocaulus magnicornis Cope, 1882, (館藏編號: NAR-168)
顎骨及牙齒化石, 年代: 二疊紀(2.8億年前), 美國俄克拉荷馬州(Oklahoma)沃里卡(Waurika)附近出土。
迷齒亞綱 Subclass: Labyrinthodontia
迷齒亞綱為兩棲類進化史上最原始的族類,其中以之魚石螈被認為是首先登陸的兩棲動物。
(1) 離片椎目 Temnospondyli
1a. 真離片椎亞目 Eutemnospondyli
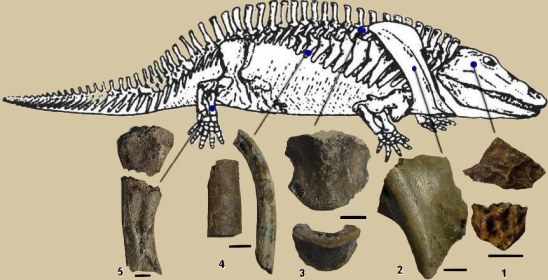
曳螈 (巨首曳螈, 中文名稱為編著所擬訂)
Eryops megacephalus Cope, 1877,
(館藏編號: NAR-056)
1.
顱骨碎片(共2片),
2.肩胛骨碎片(1片),
3. 間椎體(共2個),
4. 不完整之肋骨(共2片),
5.
下肢骨(部分已缺失),
化石於美國奧克拉荷馬州(Oklahoma)的窩利卡(Waurika)出土, 地質學層位: 二疊紀早期的萊恩形成層(Ryan
formation) 。
生活於二疊紀早期(距今約2億8千萬年前)的大型原始兩棲類,
估計體長約150厘米, 身體結構笨重, 頭部扁平而巨大, 被有堅硬的骨片, 脊椎和四肢粗壯, 半月形的椎間體(上圖3)發育較為完善, 為離片椎目成員的特徵之一。從化石臆測其復原的外形與現今的鱷相似,
但皮膚內似乎沒有堅硬的體甲片, 本種可能已經完全適應陸上生活。這群首先從海洋向陸地進發的離片椎目成員於二疊紀時期十分繁盛,
到了三疊紀時由於水生爬行類的出現以及三疊紀未期原始恐龍的繁衍使得牠們逐漸被淘汰, 最終僅有少數的品種延績至白堊紀早期, 最終完全滅絕。


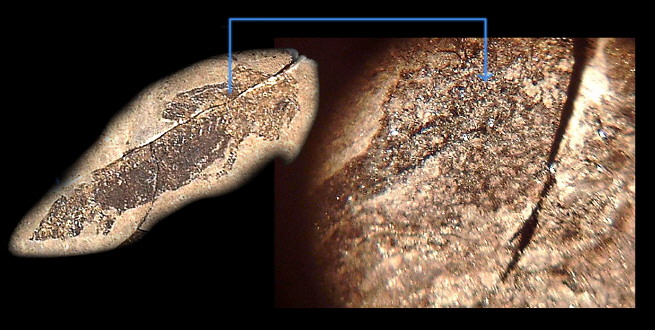
瘤狀的皮膚 (放大20倍)
異幻龍
Apateon
pedestris Meyer,1848, (館藏編號: NAR-033)
標本長5.5厘米,保存於淡水石灰岩層中,德國的Odernheim出土, 化石由德國馬得堡(Magdeburg)輸入。生活於二疊紀早期(距今約2億8千萬年前),推測牠是一種細小的完全水生兩棲類,外形酷似現今的蠑螈,牠的頭骨扁平,眼孔向上,從化石隱約可見羽毛狀的外鰓,體軀寬而扁,肢骨較纖弱,前肢上有4指,尾部為巨大的鰭。水生的離片椎類代表了兩棲動物的進化的途徑:椎體的椎間體遂漸擴大,椎側體趨向萎縮,直至整個椎體由椎間體獨立組成。於晚二疊紀至三疊紀中占優勢,但到了晚三疊紀瀕臨滅絕,白堊紀時完全消失。
滑體亞綱 Subclass: Lissamphibia
(1) 無足目 Apoda
尾極短或無, 無四肢和肢帶, 蚯蚓狀。已知160餘種, 下分隸6~7科 30 餘屬。廣布於環球熱帶和亞熱帶濕熱地區,
尤以南美洲的種類最多。非洲東西兩側均有分佈,
並在塞舌爾群島有6~7種 , 而不見於馬達加斯加島。
在亞太地區分佈于東南亞和南亞及西側新幾內亞。澳洲及歐洲無分佈。中國現有2種, 雙帶魚螈和版納魚螈,
分佈於廣西和雲南南部。
體細長, 體表周身有結紋環繞, 形成許多排環褶。每1環褶間有排列成行的長囊狀大腺體和4~6行真皮骨質小鱗, 下陷在真皮膚內,背面褶間小鱗多的達千行, 僅個別種類屬無鱗。頭側鼻與眼間有1可伸縮的"觸突", 可能與嗅覺有關。 眼小, 無眼險, 眼隱于於下或爲薄的膜骨所覆蓋。中耳僅有不發達的耳蓋骨和耳柱骨, 無咽鼓管、鼓室和鼓膜。舌較大, 有游離緣抵向內鼻孔。有上顎由和與之平行的犁腭齒各1排, 下顎有齒1~2排。 頭部背腹面的骨片均大而少, 排列緊湊而堅實, 一般無大窩孔。這種頭骨類型雖與現生的有尾類, 無尾類均不同, 而骨片成分基本相同。堅固的頭顱是適應穴居生活的一種性狀。椎體雙凹型, 有殘留脊索。肋骨較長。右肺發達, 左肺退化。雄性泄殖腔壁能翻出成爲交接器。體內受精。肛裂縱置或橫置。
除南美的盲游蚓科爲水棲外, 其餘都是穴居, 棲息在各種淡水域附近的潮濕穴洞內。夜出覓食蚯蚓、白蟻等。一般在大雨後容易見到。卵生或卵胎生。雙帶魚螈在洞穴中産卵20~50餘枚 , 以濕潤的軀體盤繞這堆卵 , 直待卵孵出。在孵出時, 3對細長羽狀外鰓已萎縮, 鰓裂沒有封閉; 幼體的極後段還有鰭褶, 有側線器官。在水中生活一段時間以後, 鰓裂封閉, 鰭褶消失, 觸突從小窩內可明顯伸出, 表示完成變態過程。卵胎生的以胚期的齒刮取取母體輸卵管壁分泌的乳汁狀物作爲營養物質。
在現生兩棲類中僅蚓螺目有骨質小鱗, 頭部骨片間一般無大窩孔; 前者是原始特徵, 後者與古兩棲類中不存在大窩的特徵相似。有關本目的起源至今尚無定論,雖曾認爲可能與殼椎亞綱的缺肢目或細螈目晚石炭紀的弛頂螈(Lysorophus)有淵源關係, 但證據卻不足。蚓螈類發展歷史悠久, 石炭紀有可疑的化石的發現。

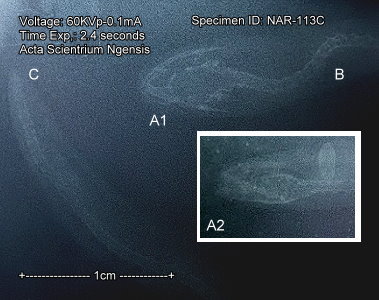
版納魚螈
Ichthyophis bannanicus Yang, 1984, (館藏編號: NAR-002113(左),
浸製標本, NAR-002113C(右) X射線攝像: A1頭部正面, A2: 頭部側面, B: 脊椎; C: 尾椎)![]()
墨西哥蛇皮蚓螈 Dermophis mexicanus (Duméril et Bibron), 1841, (館藏編號: NAR-005129,
浸製標本 )![]()

蚓 螈
Typhlonectes natans (Fischer),1879. (延陵格物研究組曾誤認為
Gymnopis 的一種,
有誤), (館藏編號:
NAR-006021, 剝製標本)![]()
標本長37.45厘米,
外表如蚯蚓或蛇, 尾短,
無四肢和帶骨。皮膚裸露, 光滑質軟,
有很多環狀皺紋, 黏液腺很發達。背部黑色,
腹部呈灰黑色略帶紫色。眼睛退化,
隱於皮下而不司視覺,
但在眼及鼻孔間的低陷部位內有收縮性觸角, 用於鑽穴之用,
聽覺器官退化, 但嗅覺卻很靈敏。營體內受精的生殖模式。廣泛分布於環球熱帶及亞熱帶地區。
(2) 有尾目 Caudata
本目成員終生有尾, 幼體與成體形態極為相似,
為現存兩棲類中較為原始的族群。全世界已知5科60餘屬350餘種。主要分布於全北界。 非洲熱帶和澳洲無有尾類。中國已知有3科15屬和35種左右。小鯢科和蠑螈科的特有屬較多。
頸部較明顯, 四肢勻稱。皮膚光滑濕潤,
緊貼皮下肌肉, 無體鱗。中耳只有耳蓋骨-耳柱骨複合結構,
無鼓膜, 水棲階段由耳肌聯接肩帶,
或由韌帶聯接鱗骨、方骨部位, 變態後或陸棲種類靠肩胛提肌的牽引傳導聲波。一般均不能發聲。舌後端不完全游離,
不能從後端翻出攝食。上下顎均有小齒,
僅鰻螈類覆以角質片。 犁骨齒排列形式不一。額骨與頂骨左右各一對,
一般沒有方軛骨, 上顎骨不與方骨相接。椎體雙凹型或後凹型,
有肋骨。肢帶軟骨質成分多, 肩帶中僅肩臼周圍的部分骨化,
左右側的喙軟骨不直接相連, 無鎖骨;
髂骨短, 恥軟骨與坐骨聯成平板狀,
有的有Y形前恥軟骨。其形體適於在水中迅速遊動,
在陸上活動時, 軀幹很少抬起離開地面,
以交替的邁步動作和軀幹與尾的波狀彎曲而移動前進。 能疾走或樹棲的種類,
其四肢較長, 或尾有攀援能力。
生活習性大多生活於淡水水域及其附近。生活在池塘、江河、湖泊、山溪、沼澤中的多爲半水棲,
其他以終生水棲或陸棲爲主。均無交接器 ;
無肛腺者爲體外受精, 有者爲體內受精,
其受精方式是: 接納雄性排出的精包,
並把精子植入泄殖腔壁的貯精囊內,
多爲卵生。在水中或水外陰濕處産卵,
卵可聚成卵帶狀、膠囊狀或單生。 個別種類卵胎生或胎生。幼體上下顎有真齒而不是角質齒;
幼體先出前肢芽, 有外鰓而無內鰓:
平衡枝的有無以及鰓和鰭褶的形狀大小,
一般與生活習性有成體保留多種幼態性狀,
如終生有鰓或有鰓裂, 或犁骨齒停滯在幼體期齒列形狀等。幼體狀態在繁殖方面即已達性成熟階段者稱爲童體型或幼態持續型,
這些終身有鰓或鰓裂的類群, 被認爲是各自獨立發生的;
最早的是三疊紀的童螈。此外, 著名
的美西螈在內外因素包括種群於不同分佈區的影響下,
也可出現童體型。
有尾目的起源主要有兩種說法: (1)直接起源於總鰭魚類扇骨魚的孔鱗魚目類(Porolepiformes)成員; (2)起源於殼椎類,
特別是石炭紀的弛頂螈(Lysorophus)。這些論點都還沒有足夠的論據。有尾目最早的化石見於早侏羅紀。
有關若干類群在變態前後,
中耳和顎部骨片以及犁骨齒,
在形態結構上有頗大的差別。鰓裂封閉是完成變態的主要標誌。
隱鰓鯢科: Cryptobranchidae

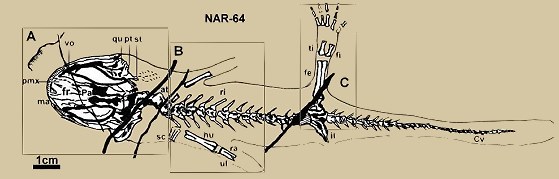
NAR-064 天義初螈 Chunerpeton tianyiensis) 線描圖部分骨骼縮寫說明
at. 寰椎(atlas),
Cv.
尾椎(Caudal vertebra),
fe.
股骨(femur),
fi.
腓骨(fibula),
fr.
額骨(frontal bone),
hu
肱骨(humerus),
il.
盤骨(ilium),
ma.
上頜骨(maxilla),
pa.
顱頂骨(parietals),
pmx.
前颌骨(premaxilla),
pt.
翼骨(pterygoid),
qu.
方形骨(quadrate),
ra.
橈骨(radius),
ri.
肋骨(rib),
sc.
肩胛喙骨(Scapulocoracoid),
st.
鐙骨(stapes),
ti.
脛骨(tibia),
ul.
尺骨(ulna),
vo.
犁骨(vomer)

天義初螈
Chunerpeton tianyiensis Gao et Shubin, 2003, (館藏編號: NAR-007064)![]()
時 代:
侏羅紀晚期~白堊紀早期
發現地點: 內蒙古赤峰市
地質層組: 道虎溝形成層
化石基質: 頁岩

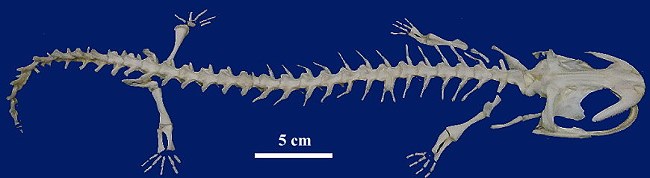
華南大鯢 Andrias
sligoi (Boulenger), 1924
[中國大鯢
Andrias davidianus (Blanchard), 1871, sensu lato] (館藏編號:
NAR-007100 幼體: 浸製標本, (上) NAR-007100A: 成體, 剝製標本, (下) 骨骼標本 NAR-007100B )![]()
現存兩棲類中最大型的物種, 因外表如魚類而且會發出如兒啼般的叫聲而俗稱娃娃魚。生活在野生環境的個體最大體長為180厘米
(NAR-007100A活飼8年標本自然死亡的個體體長60厘米), 體重可達50千克, 頭及身軀扁平, 口寬大, 上、下頜皆有細齒, 眼小,無眼瞼,
體側有明顯的與體軸平行的縱行厚膚褶及成對的疣, 體表光滑, 皮膚有腺體, 尾側扁。前、後肢短小, 前肢有4指, 後趾有5趾, 趾間有淺蹼,
擅於在石礫中爬走及游泳。生活於岩石孔洞附近水溫低於攝氏22度的水質潔淨, 水流湍急的溪澗中, 白天喜隱於暗處, 夜行肉食性,
以水生昆蟲、魚類、蝦、蟹、蛙類以及蛇類為食, 繁殖期7~9月。《國際自然保護聯盟瀕危物種紅色名錄》(IUCN
3.1) 列入極危物種, 雖然在中國進行人工繁殖的進展不錯, 但所有舉措對大鯢的保育是適得其反的: 因為最近方證明存在於中國的大鯢屬至少3~5種,
不當的混合必然引起種族的衰落, 所以廣泛所指的中國大鯢"Andrias davidianus" 己經有滅絕之虞, 外此生活於黃山一帶的種群被為是一個獨立的品種。

NAR-07100標本的頭部 X-光透視
本種為已知最知最大的現生兩棲類動物, 和傳統的中國大鯢分別在於: 本種頭部比較寬而扁平, 鼻孔間距更大, 口吻更圓, 眼睛更突出, 身體凹陷. 四肢短而粗各趾凹陷, 外面有一層皮膜, 尾部較短, 末端呈圓形, 體色呈棕褐色點綴黑色。分布於中國南部(珠江流域和南嶺山脈的區域)。

東京小鯢(東京山椒魚) Hynobius tokyoensis
Tago, 1931, (館藏編號: NAR-008101,
浸製標本 )![]()

對馬小鯢 Hynobius tsuensis Abé, 1922, (館藏編號: NAR-008119,
浸製標本 )![]()

虎斑鈍口螈 (虎斑鯢)
Ambystoma tigrinum mavortium Baird, 1850, (館藏編號:
NAR-009166,
剝製標本)![]()
墨西哥鈍口螈(美西螈)
Ambystoma mexicanum Shaw, 1789, (館藏編號:
NAR-009084,
浸製標本)![]()

剝製標本
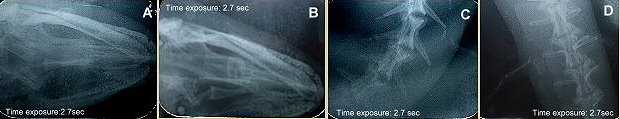
X射線對NAR-010151二趾兩棲鯢主要器官的掃描影像(剝製前): A.頭部正面; B. 頭部上45度; C.前肢;
D.後肢
二趾兩棲鯢 Amphiuma
means Garden, 1821, (館藏編號: NAR-010151,
浸製標本)
體長超過1米, 標本長72厘米。終生水中的兩棲類,
四肢極為微小, 趾的數量成為品種的重要依據。
海灣泥螈 Necturus
beyeri Viosca, 1937, (館藏編號: NAR-013139,
浸製標本)![]()


香港瘰螈的剝製標本 (經著色:
右: NAR-001T: 繁殖期雌性,
技術改善製作法)

大帽山近山頂發現的個體標本 (未作任何標本技術處理)
香港瘰螈 (香港准中螈) Paramesotriton hongkongensis
(Myers
et Leviton) 1962, [館藏編號:
NAR-015001, 高分子透明樹脂密封技術標本:
B 幼體, C 成體]![]()
異名:
Trituroides hongkongensis (Myers & Leviton, 1962)、Paramesotriton hongkongensis (Freytag, 1962)、中國瘰螈香港亞種 Paramesotriton
chinensis hongkongensis (Fei, Ye, and Huang, 1991)
體長11~14厘米, 雄螈較小,軀幹渾圓且粗壯, 頭部扁平; 背部深褐色,
腹部表面黑色綴以橙紅色斑紋, 尾下部中間有一條紅線,
直至尾端。日間喜棲息於水溪的石罅中,
祗有夜間或雨後或會遠離水源處活動。以昆蟲幼蟲、水生蠕蟲、蝌蚪甚至小魚為食。
僅分布於香港,見於港島、大嶼山、九龍山頂、大帽山及沙頭角等海拔250~500米的山溪中.由於牠和東方蠑螈(Cynops
orientalis)很相似而被混淆,現證實兩個為不同屬的物種。
2006年5月我們在大帽山的山頂(海拔880~900米)的路旁偶爾發現一個可能受天敵襲擊致死的個體(見上圖), 即使在這樣高度仍可能有香港蠑螈的分布。
根據本室的格物研究組的研習, 且總結爬行動物學者的見解,
我們將之與中國瘰螈之分別綜合如下: 香港瘰螈的體形很粗壯, 頭部特別扁平, 頭蓋上的脊突(prominent
cranial ridges)明顯, 此為鑑定本種的重要特徵之一, 背部呈褐色至深褐色,
無雜色及雜斑, 尾部有橙色污斑點, 腹部橙紅至鮮紅色的斑塊之間無連接, 尾下緣有橙至紅色帶, 但是會隨個體不同而略異, 故此腹斑不能當作是鑑定依據。身體兩側各有一條由瘰塊組成,
從頭部後方到尾部前方的橫向的瘰帶。
亦有人認為: 香港瘰螈為中國瘰螈的亞種, 但從習性而觀, 本種不同於其他同屬成員,
故此多數學者認定是一個獨立的物種, 甚至有些國內文獻聲稱發現於深圳甚至華南, 但其可靠性仍有存疑。
根據最近才出版的《中國動物志:兩棲綱(上)》(2006, p.306~310)載:香港瘰螈與中國瘰螈的區別在於本種頭體的背部皮膚光滑, 疣粒較少; 體側橫突末端部位疣粒大, 形成明顯的側棱; 尾較短, 尾鰭褶簿, 腹面橘紅色斑圓形。至於四川省生物研究所認為本種是中國瘰螈香港亞種的誤解(1977)乃因缺少標本核對之故, 這個誤認一直為費梁(1990, 1999)及葉昌媛(1993)等學者引用。 直至2005年期間費梁根據實物標本對其形態特徵進行研究及參閱龐家慶等關於瘰螈屬的研究論文(1992), 以及本種與中國瘰螈有不同的產卵期 (P. chinensis: 5~6月; P. hongkongensis: 11月至翌年2月), 遂將之重新晉為獨立物種。據載本種分布於香港及深圳。
中國瘰螈 Paramesotriton
chinensis (Gray) 1859, [館藏編號:
NAR-015046, 高分子透明樹脂密封技術標本]![]()
本種與香港瘰螈明顯不同, 瘰粒較多且密, 但與廣西瘰螈 [P. guanxiensis (Huang, Tang et Tang, 1983], 極為相似, 體上之瘰粒的大小及密度極為接近, 但本種的吻長與眼徑之長度大概相近, 廣西瘰螈的吻長大於眼徑。中國瘰螈不產於香港, 但常見於本地的水族市場。

剝製標本 (經著色)

未作任何標本技術處理, 館藏編號:
NAR-041A, 長期保存狀態: 浸製
無斑肥螈 Pachytriton labiatus (Unterstein),1930, [(館藏編號:
NAR-015041,
剝製標本(1) /
浸製標本(1)]![]()

東方蠑螈 Cynops
orientalis (David), 1873, (館藏編號: NAR-015040, 剝製標本)![]()
常見的小型蠑螈, 多見於寵物市集中, 體長一般少於6厘米, 背部深黑色, 皮膚光滑, 腹部以橙紅色為主, 綴以黑色橢圓形斑塊, 尾部呈劍形。以昆蟲及小型水生生物為主, 分布於中國中部及東部地區。

歐洲真螈(火蠑螈) Salamandra salamandra L., 1758, (館藏編號:
NAR-015069,
浸製標本)![]()
亞種: S. s. morenica (Joger
et Steinfartz), 1994
藍尾蠑螈 (楚雄蠑螈)
Cynops cyanurus Liu, Hu and Yang, 1962, (館藏編號:
NAR-055,
浸製標本)![]()
沖繩劍尾蠑螈 Cynops ensicauda popei (Inger), 1947,
(館藏編號:
NAR-015102,
浸製標本)![]()
日本紅腹蠑螈 Cynops
pyrrhogaster (Boie), 1826, (館藏編號:
NAR-015083,
浸製標本)![]()
琉球棘螈 Echinotriton andersoni (Boulenger), 1892,
(館藏編號:
NAR-015108,
浸製標本)![]()
貴州疣螈 Tylototriton kweichowensis (Fang et Chang), 1932,
(館藏編號:
NAR-015092,
剝製標本)![]()

小鰻螈 Siren
intermedia Barnes,1826, (館藏編號:
NAR-016061,
浸製標本, 活飼自然死亡)![]() 二趾兩棲鯢60厘米, 呈圓筒形, 完全水生的鰻形的有尾兩棲類, 終生保留鰓及鰓裂, 具3對外鰓, 眼和口都很細小,
沒有眼簾, 上下頜上未有齒的分化而祗有角質鞘, 犁骨終生保留幼始型結構。
尾部側扁, 極短, 僅有一對很短的前肢,有4指, 但沒有後肢。生於流速緩慢的溪流中, 主要以分昆蟲和小型無脊椎動物為食。北美洲特有物種,分布於美國(由南卡羅納州至德薩斯州和密西西比河流域至伊利諾及印第安地區) 。
二趾兩棲鯢60厘米, 呈圓筒形, 完全水生的鰻形的有尾兩棲類, 終生保留鰓及鰓裂, 具3對外鰓, 眼和口都很細小,
沒有眼簾, 上下頜上未有齒的分化而祗有角質鞘, 犁骨終生保留幼始型結構。
尾部側扁, 極短, 僅有一對很短的前肢,有4指, 但沒有後肢。生於流速緩慢的溪流中, 主要以分昆蟲和小型無脊椎動物為食。北美洲特有物種,分布於美國(由南卡羅納州至德薩斯州和密西西比河流域至伊利諾及印第安地區) 。
鰻螈亞目在分類系統中的位置仍有爭議, 故此把它放在有尾目的末段討論, 編者不認為本亞目成員在兩棲類進化中代表較先進的族群, 反而猜測其位置是介於隠鰓觬亞目和蠑螈亞目之間。
(3) 無尾目 Anura
兩棲類中最大的一目, 亦為最高等的族群, 現存20多科, 300多屬3500餘種。除南極外, 廣布於各大洲;
在溫差小, 濕度大的中美、南美和非洲熱帶和亞熱帶種類最多,
個別達北極圈南緣。中國已知近230 種, 分隸7科25屬。
體體寬而短, 後肢長, 跗部自成一節, 適 於跳躍或游泳。皮膚一般光滑濕潤, 有的皮膚上有角質刺, 或頭頂部皮膚骨質化而全無小鱗, 皮膚與皮下肌肉之間有一大淋巴囊。 口大, 舌後端多游離,可翻出攝食, 下眼簾連有透明的瞬膜。眼大, 位於頭側, 突出時可擴大視野。下顎無齒, 上顎一般有細齒。中耳多完備, 鼓膜顯著或隱于皮下或無。雄蛙一般有共鳴器(聲囊) , 這是一種第二性徵。 額骨與頂骨癒合爲額頂骨, 有方軛骨。椎體有雙凹型、後凹型、變凹型、前凹型或參差型。椎骨一般爲10 枚, 薦椎後爲一細長的尾杆骨。腰帶呈"U"字形, 髂骨長, 平直向前與薦推橫突相關聯。脊柱短而數少, 彎曲度和靈活性不強, 前三對軀椎上或有短肋, 為原始性狀, 見於低等類群。脊柱與後肢的關聯由髂骨、後肢骨和尾杆骨部位的肌肉、肌腱而加強, 可以作有力的攀援、跳躍或對稱蛙式游泳動作。前肢較強, 主要可減輕落地時的衝擊力。肩帶左右側上喙骨或相接(固胸型)或相互重疊(弧胸型) , 或有弧固型。橈骨與尺骨、脛骨與腓骨分別癒合爲橈尺骨和脛腓骨; 近端2跗骨長, 自成1 節, 增強了跳躍、游泳能力。無尾類體形體現了與運動行爲相適應的結構。多數種類大多蹲在以後肢向後掘的淺泥凹內 , 既可防失水, 又能坐待捕食 , 捕食時後肢仍貼於地面。
無交接器, 體外受精。卵生 , 僅個別種卵胎生或胎生。 幼體 ~ 蝌蚪的體形、食性等與成性迥然不同。蝌蚪口部或有角質小齒; 有鰓和尾, 早期爲外鰓, 不久外鰓萎縮, 代之以內鰓, 舌弓部位皮膚逐漸向後生長, 成爲覆蓋著內鰓的鰓蓋褶, 再與腹位的皮膚癒合, 形成鰓蓋腔 , 僅留有1或2個小出水孔, 出水孔或位腹面中部或兩側或位體左側 , 出水孔通向體外 , 依據角齒之有無和出水孔之特徵, 分爲 4 個類型: (1)角齒腹孔型; (2)角齒左孔型; (3)無角齒腹孔型及 (4)無角齒雙孔型。口部一般有成行的角質小齒和角質顎。以刮取藻類爲主, 無者以吞食浮游生物爲主。蝌蚪先出後肢芽, 前肢於早期就在鰓蓋腔內或附近部位發育, 變態高潮時, 前肢伸出體外, 尾、鰓萎縮以至消失, 器官改組, 呼吸和消化器官尤爲突出。此時一般不攝食, 也不能久潛于水中 , 否則大多要溺死。
本目動物有水棲、半水棲、陸棲、樹、穴居等不同的種類。一般棲息於各種淡水水域及其附近潮濕的林木草叢中。在黃昏、黎明或雨後的白天活動頻繁, 覓食各類昆蟲及其幼蟲和蠕蟲等。繁殖習性多樣化, 有以各種方式撫育卵或幼體的種類。
發現於馬達加斯加的早三疊世化石三疊尾蛙, 曾稱原蛙, 有與近代型無尾目基本相似的主要特徵。一般認爲它是無尾類的先驅, 形態上已具備向典型無尾類演化發展的基礎。近代型的於阿根廷發現的比較完整的侏羅紀化石,
以色列曾發現有許多完整的早自堊紀化石無舌類。 可見幾個主要原始類群都已出現於中生代未期,
而且分化速度相當快。一般認爲雙凹型、雙凹型無尾兩棲類屬於原始類群, 而後凹型及變凹型無尾兩棲類屬於高級類群; 鋤足蟾科被認爲是介於原始型的和高級型的中間類群。
有尾目和無尾目的過渡體雖然相像, 但彼此在肩帶類型, 椎體類型以及外部形態特徵等皆大為相異, 如無尾目成員有嗚囊, 而牠們的舌部, 牙齒結構及皮膚的結構亦不同, 此等皆為分類的依據之一。
3a. 始蛙亞目 Archaeobatrachia
鈴蟾科: Bombinatoridae
東方鈴蟾 Bombina
orientalis (Boulenger), 1890, (館藏編號:
NAR-018008, 剝製標本, 見總圖)![]()
3b. 中蛙亞目 Mesobatrachia
劉氏掌突蟾
Leptolalax laui Sung et al., 2014, (館藏編號: NAR-022131)![]()

短足角蟾 Xenophrys
brachykolos (Inger et Romer), 1961, (館藏編號:
NAR-022053 , 蝌蚪期、蛙期 浸製標本)![]()
雷山髭蟾
Vibrissaphora leishanensis (Liu et Hu), 1973, (館藏編號: NAR-022117 ,
蛙期 浸製標本)![]()
北美鋤足蟾科: Scaphiopodidae
庫氏掘足蟾 Scaphiopus
couchii Baird, 1854, (館藏編號: NAR-022117 ,
幼蛙期 浸製標本)![]()

負子蟾(蘇利南爪蟾) Pipa pipa
L., 1758, (館藏編號:
NAR-026067)![]()
3c. 新蛙亞目 Neobatrachia
黑眶蟾蜍 Bufo malanostictus Schneider
1799, (館藏編號: NAR-031006,
剝製標本, 見總圖)![]()
玻璃蛙
Hyalinobatrachium (Centrolenella) valerioi Dunn, 1931, (館藏編號:
NAR-031006, 剝製標本)![]()
蘇利南角蛙
Ceratophrys cornuta (L.), 1758, (館藏編號:
NAR-034091, 剝製標本)![]()
秘魯角蛙 Ceratophrys
stolzmanni Steindachner, 1882, (館藏編號:
NAR-034098, 剝製標本)![]()

金色箭毒蛙
Phyllobates terribilis Myers, Daly et Malkin, 1978, (館藏編號: NAR-035106,
浸製標本)![]()
鈷藍箭毒蛙
Dendrobates tinctorius azureus (Hoogmoed), 1969, (館藏編號: NAR-035156,
浸製標本)![]()
迷彩箭毒蛙
Dendrobates auratus (Girard), 1855, (館藏編號: NAR-035107,
浸製標本)![]()
卵齒蟾科: Eleutherodactylidae

溫室蟾
Eleutherodactylus planirostris (Cope), 1862, (館藏編號: NAR-036187,
浸製標本)![]()
隨園藝植土引入香港的特殊蛙類, 已經在野生環境中成功繁衍,
因習性和體型跟盧文氏樹蛙 (Philautus romeri Smith, 1953)相似而被人以保護本地原生生物而被刻意捕捉,
但似乎成效不彰。常見於雨後各區的公園中的泥圃中。標本採自香港島歌賦山。
紅眼樹蛙 Agalychnis
callidryas Cope, 1862, (館藏編號: NAR-039118,
浸製標本)![]()
中國雨蛙 Hyla
chinensis (Gunther),1859
, [館藏編號:
NAR-039017, 剝製標本(1), 見總圖; 及剝製標本(1)]![]()
亞馬遜牛奶蛙
Trachycephalus resinifictrix (Goeldi), 1907, (館藏編號:
NAR-039093, 剝製標本)![]()
納塔爾葉折疊蛙
Afrixalus spinifrons (Cope), 1862, (館藏編號: NAR-041147, 剝製標本)![]()
金色曼蛙 Mantella
aurantiaca Mocquard, 1900, (館藏編號:
NAR-043114, 浸製標本)![]()
馬島曼蛙 Mantella
madagascariensis Grandidier, 1872, (館藏編號: NAR-043136, 浸製標本)![]()
亞洲錦蛙 Kaloula
pulchra pluchra Gary
1831, (館藏編號: NAR-044005, 剝製標本,
見總圖)![]()
飾紋姬蛙 Microhyla
ornata (Dumeril et Bibron),
1841, (館藏編號: NAR-044009, 剝製標本,
見總圖)![]()
花姬蛙 Microhyla
pulchra (Hallowell), 1861, (館藏編號: NAR-044051,
浸製標本)![]()
● 蛙亞科: Raninae
貢德氏蛙 Rana gutheri Boulenger
1882, (館藏編號: NAR-047004 / NAR-047004A , 浸製標本)![]()
體長約10厘米,
呈棕色或褐紅色, 有兩條金黃色或帶褐紅色的縱線,
由背側下延, 唇部有白色斑點,
腹部白色, 皮膚平滑,
腹側黃白色, 有棕色小斑,後肢有深色間條。本種廣泛分布於農地、池塘或塹壕。棲於平緩的水流中,
以蟋蟀、蚱蜢及地棲蜚蠊及蚯蚓為食; 有時亦捕食其他蛙類。有毒腺,
但毒性不強。本目為兩棲綱中數量最多,
體形最為特殊的類群, 現有20多科約3500種,
一般體闊而短, 後肢很長,
跗部僅一節, 皮膚一般光滑,但也有的具角質刺,
眼部及口部大, 舌部可翻出攝食。水棲、半水棲、陸棲或樹棲以至穴居動物,擅跳。
古氏赤蛙(大頭蛙)
Limnonectes kuhlii (Tschudi), 1838 (= Rana kuhlii
Tschudi, 1838), (館藏編號: NAR-047054 ,浸製標本)![]()
標本採自台灣(台北烏來)。
虎紋蛙 Rana tigrina sp. rugulose Wiegumann
1834, (館藏編號: NAR-047002,
剝製標本, 見總圖)![]()
大綠蛙 Rana livida
(Blyth), 1856, (館藏編號:
NAR-047052 ,浸製標本)![]()
闊褶水蛙 Hylarana (Hylarana) lutouchii (Boulenger), 1899, (館藏編號: NAR-047049 , 幼體浸製標本)

香港湍蛙
Amolops hongkongensis (Pope et Romer), 1951, (館藏編號:
NAR-047050A 卵, B:成體, 浸製標本, 圖示 NAR-047050: 剝製標本, 復原色)![]()
叉舌蛙科: Dicroglossidae
澤 蛙 Fejervarya
limnocharis (Gravenhorst), 1829, (館藏編號:
NAR-047A045 ,浸製標本)![]()
福建大頭蛙
Limnonectes fujianensis Ye et Fei, 1994, (館藏編號: NAR-047A182 ,浸製標本)![]()
尖舌浮蛙 Occidozyga
lima (Gravenhorst), 1829, (館藏編號:
NAR-047A179 ,浸製標本)![]()

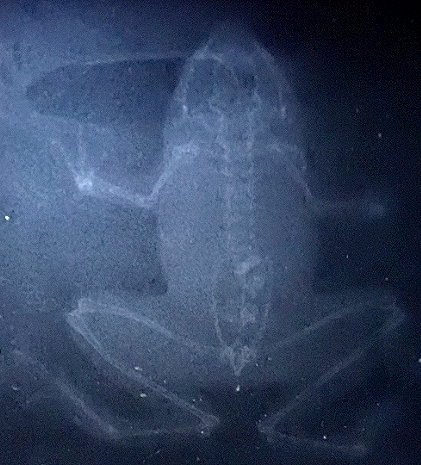
盧文氏樹蛙(盧氏小樹蛙)
Philautus romeri Smith, 1953, (館藏編號:
NAR-049043, 浸製標本, 右圖: 骨骼系統的X-光透視)![]()
香港獨有品種,
僅零散地分布於馬灣海峽至東博寮海峽線以西至南部島嶼中的大嶼山、赤鱲角、南丫島及蒲台島,其分布特點頗有研究意義。但是由於赤鱲角新機場的建立,該地區的族群自1991年開始被人不自然地引入香港島及九龍和新界至少七個地點進行遷地保育。奇怪的是,
經過十多載繁衍歲月的今天, 我們從未有盧文氏樹蛙的非自然棲地有任何個體的紀錄(10
/ 2006), 反而在其原生地有較密集蝌蚪群落的發現。盧文氏樹蛙無疑是舉世聞名的香港奇異物種,
保育本種為香港大自然愛護者的共同責任。
華萊士飛蛙(黑掌樹蛙) Rhacophorus nigropalmatus Boulenger,1895, (館藏編號:
NAR-049088, 剝製標本)![]()
台灣褐樹蛙 Buergeria robusta (Boulenger),
1909, (館藏編號:
NAR-049060, 浸製標本)![]()
斑腿泛樹蛙 Polypedates megacephalus
Hallowell, 1961, (館藏編號:
NAR-049044, 浸製標本)![]()
屏邊泛樹蛙 Polypedates pingbianensis Kou et
al., 2001, (館藏編號: NAR-049148, 浸製標本)![]()
棕褶樹蛙 Rhacophorus feae (Boulenger),
1893, (館藏編號: NAR-049149, 浸製標本)![]()
 |
本網頁中所有圖片及文本受延陵科學綜合室版權保障
All images and text on this page are
copyright protected © Acta Scientrium Ngensis